

Abstract
Introduction: Using whole blood transcriptional profiling, we have previously shown deregulation of several interferon-stimulated genes (ISG) and deregulation of immune and inflammation genes in MPNs as well. Most lately, we have shown that deregulated HLA-genes are upregulated by interferon-alpha2 (IFN) treatment. Herein, we for the first time describe the landscape of ISGs during treatment with IFN, the aims being to describe if unique ISG transcriptional signatures might be elicited during IFN treatment with potential differences between subgroups.
Methods: Eight patients with ET, 21 patients with PV, and 4 patients with PMF participated in the study. All patients received treatment with IFN, in the large majority in a dosage ranging from 45-90 ug x 1 sc/week. Gene expression microarray analysis of whole blood was performed before and after 3 months of treatment. Total RNA was purified from whole blood, amplified to biotin-labeled RNA, and hybridized to Affymetrix HG-U133 2.0 Plus chips recognizing 54,675 probe sets (38,500 genes).
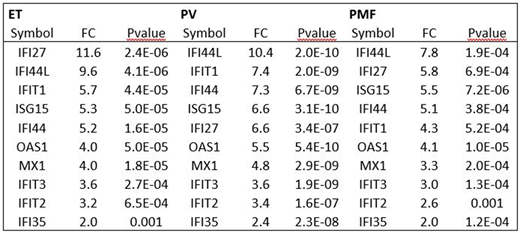
Results: We identified 6261, 10,008, and 2828 probe sets to be significantly differentially expressed in ET, PV, and PMF, respectively, in response to treatment with IFN (pvalue < 0.05). Twenty-one previously identified ISGs were investigated: Six genes involved the response to virus: ISG15, IFI35, IFI44, IFIH1, MX1, and OAS1, 2 transcriptional regulators: IFI16 and SP110, and 13 IFN-inducible genes: ADAR, CXCL10, IFI6, IFI27, IFI30, IFI44L, IFIT1, IFIT2, IFIT3, IFITM1, IFITM2, PSME1, and PYHIN1. Significant upregulation of 20, 21, and 18 ISGs was found in patients with ET, PV, and PMF, respectively. None of the 21 genes was significantly downregulated in any of the patient groups. The 6 genes involved in response to virus and the 2 transcriptional genes were significantly upregulated in all subgroups. In patients with ET all but one gene (CXCL10) of the 13 IFN-inducible genes were significantly upregulated. In patients with PV all 13 IFN-inducible genes were significantly upregulated and in PMF all but three (CXCL10, IFITM2, PUHIN1) IFN-inducible genes were significantly upregulated. No significant differences between subgroups were recorded (data not shown).
Discussion and Conclusions: Eighteen genes were significantly upregulated compared to baseline values in all 3 MPN-subgroups. The CXCL10 gene was not significantly regulated in ET and PMF during IFN treatment but significantly upregulated in PV. CXCL10 is involved in the activation of neutrophils. Accordingly, impaired regulation of CXCL10 on exposure to IFN might compromise recruitment and activation of neutrophils. The excessively high expression levels of IFI27 after IFN are intriguing but might be explained by the reported high circulating levels of regulatory T cells (Tregs) after IFN. Thus, Tregs produce TGF-beta which stimulates IFI27. Indeed, TGF-beta was significantly upregulated in ET and PV but not in PMF, likely due to the low number of PMF patients. Surprisingly, we did not find any differences in ISG responses between MPNs. Indeed, taken into account that the MPNs depict a biological continuum from the early cancer stages (ET/PV) to the advanced "metastatic" myelofibrosis stage, different ISG signatures between the subtypes might be anticipated for several reasons. First, such differentiated signatures might exist merely reflecting the heterogeneity between subgroups in regard to "tumor burden". Second, differences in frequencies and functionality of immune cells between the subgroups as shown in most recent studies might impact the ISG signatures. Third, chronic inflammation, considered the driving force for clonal evolution in MPNs, has been shown to compromise cell responses to IFN, implying that PMF patients with the most pronounced inflammatory state might exhibit blunted ISG responses. However, the low number of PMF patients may account for the non-significant difference between ET/PV and PMF. In conclusion, except for 4 genes (CXCL10, IFITM1, IFITM2, and PYHIN1), our study has shown the ISGs to be significantly upregulated in all MPN- subtypes after 3 months of exposure to IFN. The reasons for the absence of upregulation of the above 4 genes are unknown but chronic inflammation and immune deregulation might be influential. Further transcriptional studies of ISGs during IFN treatment in larger study populations and with longer exposure times are needed to address these issues.
Hasselbalch:Novartis: Research Funding.

This feature is available to Subscribers Only
Sign In or Create an Account Close Modal